- 1 CHAPITRE 2 : LA SEXUALITE DU MAIS, LA TRANSMISSION DES GENES D’UNE GENERATION A L’AUTRE ET LE BRASSAGE GENETIQUE
- 2 Resume
- 3 La sexualite du mais
- 4 Le genotype du mais
- 5 Transmission du genome lors de la gametogenese et la fecondation
- 6 De la composition allelique au phenotype : l’expression des genes
- 7 Conclusion
- 8 Source
- 9 Document realise dans le cadre du projet COVALIENCE
CHAPITRE PRECEDENT : CHAPITRE 1 : ROLE ET FONCTIONNEMENT BIOLOGIQUE DE L’EXPRESSION DES GENES : Comment une mouche ou un roseau peuvent-ils emerger d’un code a 4 lettres: l’ADN ?
1 CHAPITRE 2 : LA SEXUALITE DU MAIS, LA TRANSMISSION DES GENES D’UNE GENERATION A L’AUTRE ET LE BRASSAGE GENETIQUE
2 Resume
Ce chapitre présente la sexualité du maïs, la structure de son génome et comment s’opère le brassage génétique lors de la formation des gamètes et de la fécondation.
Certaines notions définies dans le chapitre 1 sont utiles pour comprendre le chapitre 2 :
Dans le chapitre 1 : «Rôle et fonctionnement biologique de l’expression des gènes», nous avons défini le gène comme étant le plan de fabrication d’un organisme vivant. Cette information est contenue dans le noyau de chaque cellule de l’organisme sous la forme d’un long code de 4 lettres (G, A, T, C). Nous avons aussi décrit comment, par différents mécanismes physiologiques, l’information génétique pouvait engendrer un phénotype. Le phénotype, c’est tout ce que l’on peut observer d’un être vivant.
Cependant, toutes les caractéristiques du phénotype ne s’expliquent pas uniquement par son origine génétique mais aussi par l’environnement dans lequel évolue l’individu.
2.1 Pourquoi lire ce chapitre
Dans cet article nous allons étudier comment se transmet l’information génétique d’une génération à une autre. Par les mécanismes de la production des gamètes et de la fécondation nous verrons comment le brassage génétique permet d’engendrer des individus sans cesse différents. En sélection paysanne, nous choisissons les plantes qui constitueront la génération suivante. Connaître les phénomènes de transmissibilité des caractères et d’expression des gènes permet de mieux comprendre ce que l’on observe et d’effectuer des choix davantage éclairés lors de la sélection.
3 La sexualite du mais
3.1 Les appareils reproducteurs
Le maïs est une espèce qui porte sur le même pied des organes reproducteurs mâles et des organes reproducteurs femelles, on dit que c’est une plante « monoïque ». À l’inverse, les plantes « dioïques », comme le houx ou le kiwi par exemple, présentent soit des spécimens mâles, soit des spécimens femelles.
Les organes reproducteurs mâles se développent au sommet de la plante et sont portés par ce qu’on appelle la panicule. La panicule présente un brin central dressé puis des nombreuses ramifications qui ressemblent un peu à des très longs épis de blé. La panicule est constituée d’une multitude d’épillets regroupant deux glumes (sorte de petite feuille) et deux fleurs mâles. Chaque fleur mâle possède 3 « sacs de pollens » que l’on nomme les anthères. Au moment de la floraison, les épillets s’ouvrent et laissent pendre les anthères qui libèrent alors le pollen dans l’atmosphère.

Figure 3.1: Panicule de maïs et zoom sur les anthères sorties des glumes et prêtes à s’ouvrir pour libérer le pollen
Les organes reproducteurs femelles sont situés plus bas sur la plante. Ils sortent progressivement d’un petit cornet de feuilles, appelées « spathes », au milieu de la plante sous la forme de longs filaments : les « styles », mais que l’on nomme plus communément « les soies ». Ces soies sont blondes ou pourpres selon les plantes et peuvent mesurer jusqu’à 40cm. À chaque soie correspondra un futur grain de maïs si celle-ci parvient à capter un grain de pollen.

Figure 3.2: Illustrations des soies du maïs
Avec cette répartition particulière des fleurs, le maïs est ainsi une espèce dite « allogame » car les plantes sont en pollinisation ouverte : c’est-à-dire que les styles (les soies) d’un individu peuvent-être fécondés par le pollen d’un autre individu. C’est le cas par exemple du maïs, du tournesol, des courges,… Mais d’autres espèces sont autogames : leurs fleurs sont protégées et s’autofécondent, il n’y a donc pas de croisement entre les individus d’un peuplement végétal de la même espèce. C’est le cas des céréales à paille (blé, orge, avoine,..). Il y a cependant parfois un petit peu d’allogamie chez les plantes autogames et souvent de l’autofécondation chez les plantes allogames.
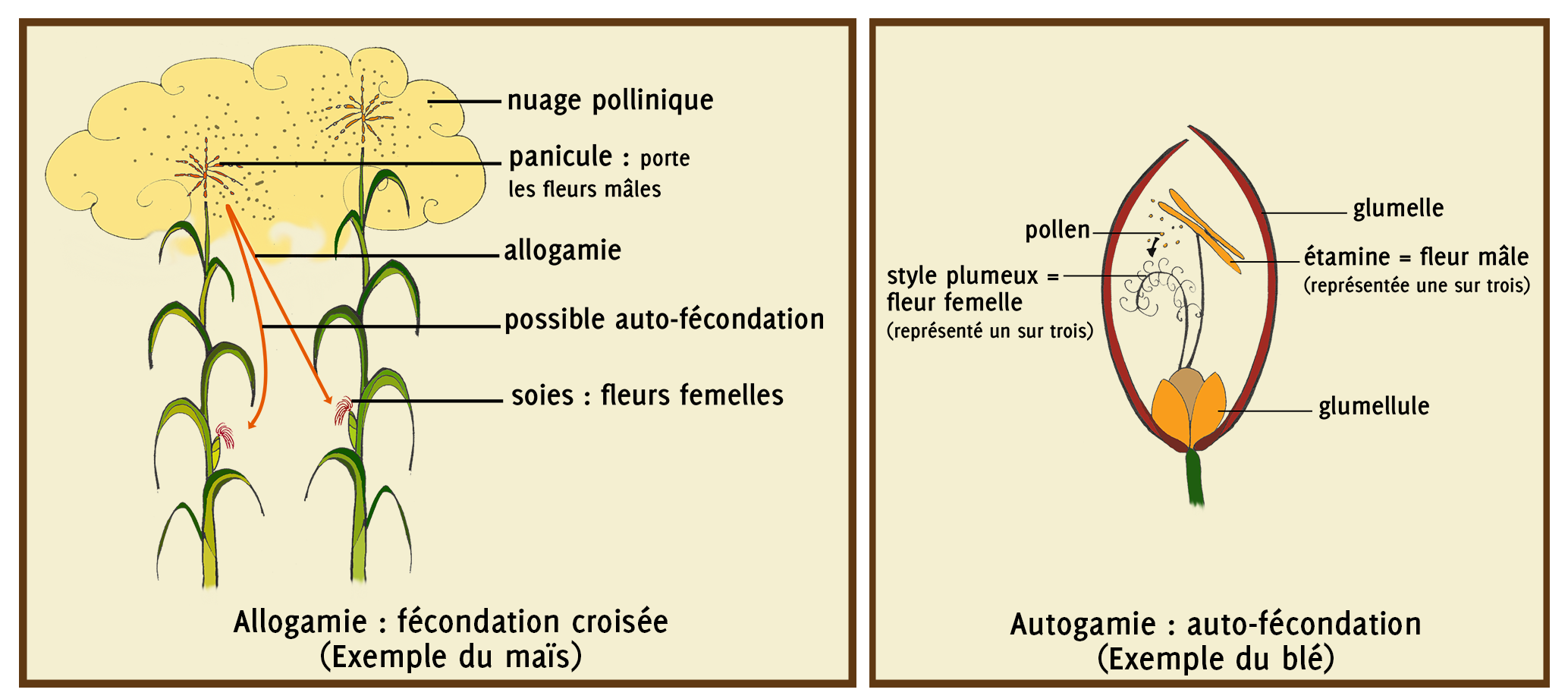
Figure 3.3: Illustration du fonctionnement de l’allogamie et de l’autogamie
3.2 La formation des gametes : la gametogenese
Pour engendrer un nouvel être vivant, les parents doivent d’abord produire des gamètes qui fusionneront pour former un embryon. Chez les humains, les gamètes mâles sont appelés spermatozoïdes et les gamètes femelles ovules. Chez les plantes, le gamète mâle n’a pas de nom particulier et on appelle le gamète femelle l’oosphère.
Les gamètes sont des cellules particulières car elles ne portent que la moitié de l’information génétique de l’organisme qui les produit.
Les gamètes sont produits dans des tissus particuliers de l’organisme (ovaires et testicules pour les gamètes femelles et mâles chez les humains, dans le sac embryonnaire et le grain de pollen chez les plantes). Voici le déroulement des divisions cellulaires qui amènent à la production des gamètes.
3.2.1 Ce qui se passe pour l’organe de reproduction femelle :
La formation du gamète femelle commence à partir d’une cellule « normale » qui possède toute l’information génétique de la plante nommée « cellule sporogène ». Cette cellule sporogène se divise en deux nouvelles cellules. Puis il y a une nouvelle division, on passe alors de 2 à 4 cellules : les macrospores. Lors de ces divisions, l’information génétique est fragmentée, chaque macrospore contient environ la moitié de l’information génétique de la plante.
Ces divisions de cellule qui amènent à la fragmentation de l’information génétique sont nommées des « méioses ».
Sur les 4 macrospores créées par la plante, seule l’une d’entre-elles va survivre, les autres vont avorter. Cette cellule fertile va alors subir une série de duplications, des « copier-coller » appelés « mitoses », qui mèneront à la formation de 8 cellules identiques sur le plan génétique contenues dans le sac embryonnaire. Le gamète femelle est l’une de ces cellules, l’oosphère. Deux autres cellules vont aussi nous intéresser pour la suite : il s’agit des noyaux polaires.
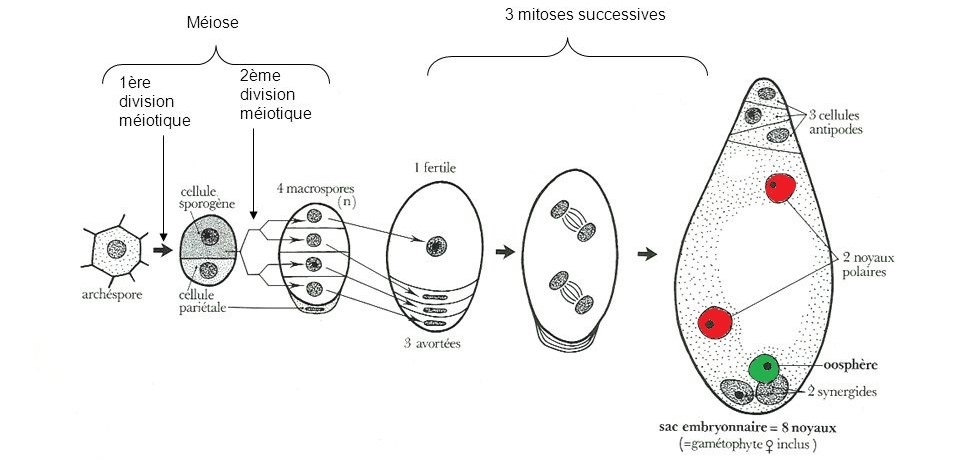
Figure 3.4: Schéma de la formation du gamète femelle : l’oosphère
3.2.2 Ce qui se passe pour l’organe de reproduction mâle :
La formation du gamète mâle, qui viendra féconder l’oosphère, résulte d’une série de transformations biologiques appelée spermatogenèse. Ce processus biologique est presque généralisable à toutes les espèces de plantes. La spermatogenèse débute dans les anthères (sac à pollen accrochés à la panicule, au sommet du pied de maïs) avec une cellule nommée le « microsporocyte » dont le noyau porte toute l’information génétique de l’individu qui produit le pollen. Cette cellule va subir 2 divisions successives qui viennent fragmenter l’information génétique ; nous verrons plus loin dans l’article le détail de cette fragmentation au niveau du génotype.
4 petites cellules, qui possèdent chacune la moitié de l’information génétique de la plante productrice de pollen, vont progressivement mûrir et devenir des grains de pollens. Le grain de pollen, à l’image du spermatozoïde, est le vaisseau de l’information génétique que l’on peut considérer comme « paternelle ». Il existe alors chez les plantes une particularité que l’on ne trouve pas chez les animaux : le noyau qui encapsule la moitié de l’information génétique paternelle va se dupliquer, c’est un « copier-coller » qui produit alors 2 gamètes mâles identiques dont nous verrons le devenir juste après.
Le grain de pollen est libéré dans l’atmosphère pas les anthères et viendra, si la chance lui sourit, se déposer sur une soie de maïs qui est couverte de tout petits poils pour favoriser l’interception des pollens. Une valse chimique fine et complexe va se lancer entre la soie et le pollen et si la compatibilité est « bonne », alors le pollen sera autorisé à germer dans la soie afin de pouvoir déposer les gamètes mâles au niveau du sac embryonnaire situé à l’extrémité de la soie grâce à son tube pollinique.
![Schéma de la formation du pollen et du gamète mâle [1]](../../../../../img/publications/chap2/formationpollen.jpg)
Figure 3.5: Schéma de la formation du pollen et du gamète mâle [1]
3.3 La fecondation
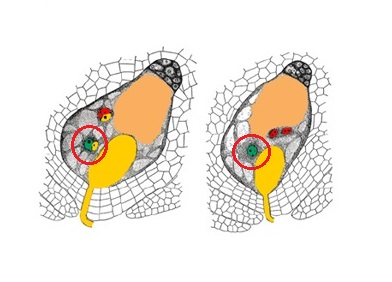
Le tube pollinique progresse rapidement dans la soie pour atteindre le sac embryonnaire qu’il va alors pénétrer pour libérer ses deux gamètes. Il s’opère alors ce qu’on appelle la double fécondation :
l’une des gamètes mâle va venir fusionner avec l’oosphère pour donner une cellule souche avec une information génétique « complète » car l’oosphère contenait la moitié de la génétique de la plante qui a donné la fleur femelle et le gamète mâle contenait la moitié de l’information de l’individu qui produit le pollen. C’est la première fécondation. La cellule souche va se diviser pour former l’embryon du grain de maïs qui germera alors plus tard pour donner une nouvelle plante.
le second gamète mâle va fusionner avec les deux noyaux polaires et donner une cellule bien particulière car elle portera deux moitiés identiques de l’information génétique de la plante mère et une moitié de l’information génétique de l’individu qui a produit le pollen. Cette cellule est donc triploïde car elle contient 3 chromosomes par type de chromosome. Elle va se diviser pour former la chair du grain appelé endosperme. C’est la seconde fécondation.
![Schéma de la double fécondation chez le maïs [2]](../../../../../img/publications/chap2/fecondationmais.jpg)
Figure 3.6: Schéma de la double fécondation chez le maïs [2]
4 Le genotype du mais
Dans le chapitre 1, nous avons vu que l’information génétique, appelée « génotype », était contenue dans le noyau de toutes les cellules d’un être vivant sous la forme d’ADN et nous savons que cet ADN joue un grand rôle dans la construction du phénotype.
Nous allons désormais observer plus précisément le génotype du maïs, c’est-à-dire la manière dont est organisé l’ADN de cette espèce dans le noyau des cellules afin de comprendre comment cette information peut se transmettre d’une génération à l’autre grâce à la gamétogenèse et à la fécondation.
Le génome du maïs est constitué de 10 paires de chromosomes (chaque paire est numérotée de 1 à 10). Au sein de chaque paire, il y a une version paternelle (issue du pollen) du chromosome et une version maternelle (issue de l’oosphère). Comme il n’existe que 2 versions de chromosomes par type de chromosome, on dit que le maïs est une espèce au génome diploïde ; l’humain est aussi une espèce au génome diploïde (mais nous avons 23 paires de chromosomes). Il existe des espèces dont le génome est triploïde (3 versions de chromosomes) comme le blé dur, voire hexaploïde (6 versions de chromosomes) comme le blé tendre.
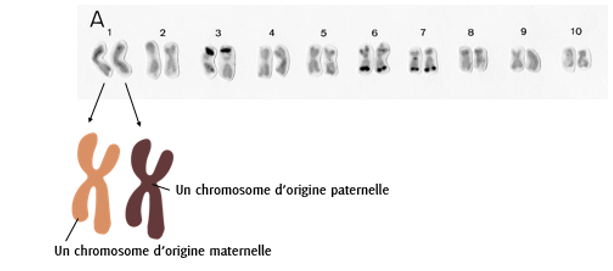
Figure 4.1: Structure du génome du maïs
Chaque paire de chromosomes porte des informations génétiques particulières. Par exemple, de façon simplifiée, le gène qui code pour la couleur du grain pourrait se situer sur les chromosomes 1, celui qui code pour la date de floraison se situer sur les chromosomes 8, etc.
Même si la réalité est plus complexe car c’est souvent le concours de nombreux gènes qui permet l’expression d’un caractère, prenons l’exemple d’un caractère codé par un seul gène, cela pourrait être le cas de la couleur de l’épi. Cet exemple va nous permettre d’introduire de nouvelles notions clefs en génétique.
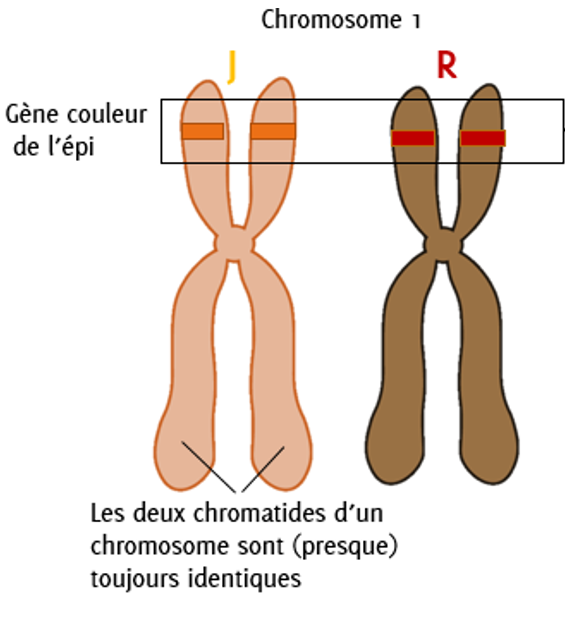
Figure 4.2: Paire de chromosome n°1
Ici, chaque chromosome porte une information différente, une version de gène différente. On appelle ces versions de gène des allèles. On a donc ici un allèle qui porte l’information « épi-jaune » et un allèle qui porte l’information « épi-rouge ».
Quand les chromosomes d’une même paire portent des versions d’allèle différentes, on dit que l’organisme est « hétérozygote » sur le gène en question » (encore un mot de vocabulaire en plus !).
Si les chromosomes portent les mêmes allèles (rouge et rouge ou jaune et jaune), alors on dit que l’organisme est « homozygote » sur ce gène. Vous vous posez peut-être la question ici « mais alors, vu qu’il y a un allèle pour la couleur jaune et un allèle pour la couleur rouge… quelle sera finalement la couleur de l’épi de ce pied de maïs ?? », patience ! Nous allons voir ceci un peu plus loin.
5 Transmission du genome lors de la gametogenese et la fecondation
Dans la partie 1 de ce chapitre, nous avons vu les différentes étapes biologiques qui permettent aux plantes de transmettre leur génétique grâce à la formation des gamètes puis par la fécondation où les gamètes paternelles et maternelles fusionnent pour donner l’embryon d’une nouvelle future plante. Dans la partie 2, nous avons observé la structure du génome du maïs, présent dans le noyau de toutes les cellules de l’organisme. Nous allons désormais observer comment le génome se fragmente, se répartit et se fusionne lors des gamétogenèses et de la fécondation.
Pour cela, nous allons d’abord présenter un modèle à une paire de chromosome, c’est-à-dire que nous considérons qu’il n’existe qu’une paire dans le génome du maïs (au lieu de 10) puis nous généraliserons plus tard à plusieurs paires de chromosomes.
5.1 Formation des gametes femelles
Dans notre exemple, la plante qui va produire les soies porte dans son génome la paire de chromosomes n°1 sur laquelle on trouve le gène de la couleur de l’épi. Cette plante est ici hétérozygote sur le gène de la couleur, son génome est [jaune – rouge].
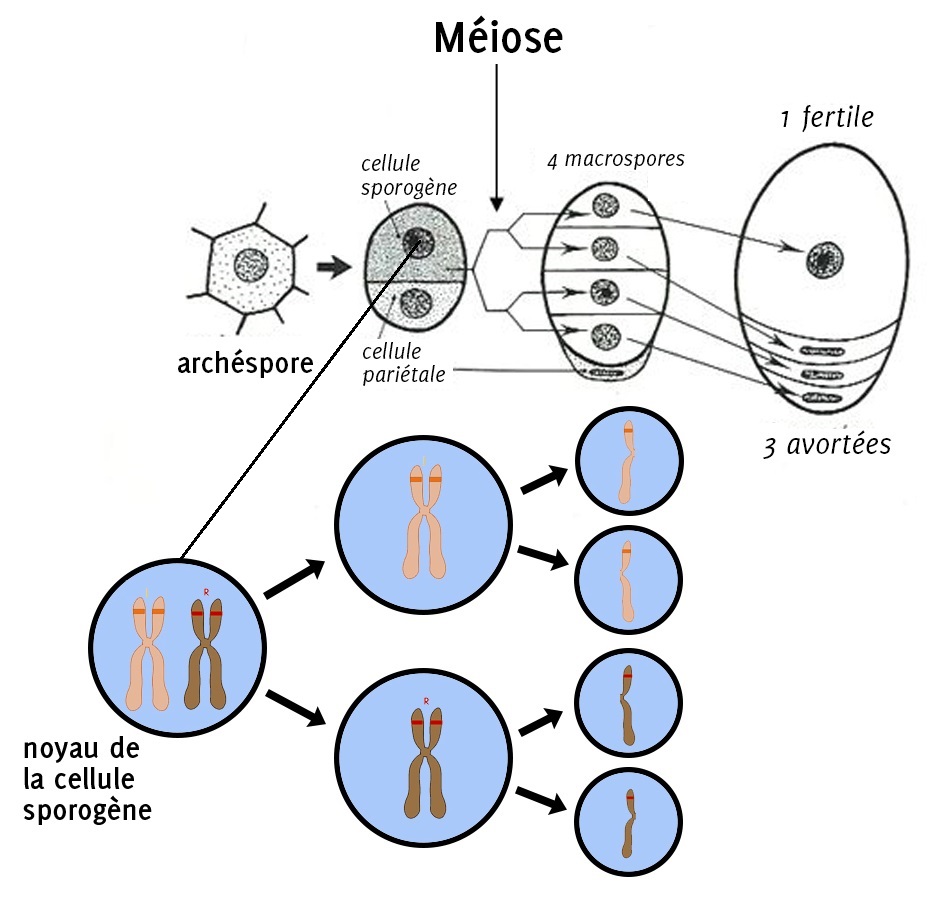
Figure 5.1: Répartition de l’information génétique lors de la méiose
Le noyau de la cellule sporogène porte le même génome que la plante dans son noyau. Lors de la méiose, cette cellule va se diviser en deux, chacune des nouvelles cellules embarque un chromosome de la paire. Au moment de la seconde division cellulaire, les chromosomes sont coupés en deux, on obtient dont 4 cellules (les macrospores) qui contiennent dans leur noyau une seule chromatide (un des deux bâtonnets qui composent le chromosome). Seulement une seule des macrospores va survivre : dans notre exemple, il y a ici 2 chances sur 4 (donc 1 chance sur 2) pour que la macrospore survivante porte une chromatide avec l’allèle [rouge] et 2 chances sur 4 pour qu’elle porte une chromatide avec l’allèle [jaune].
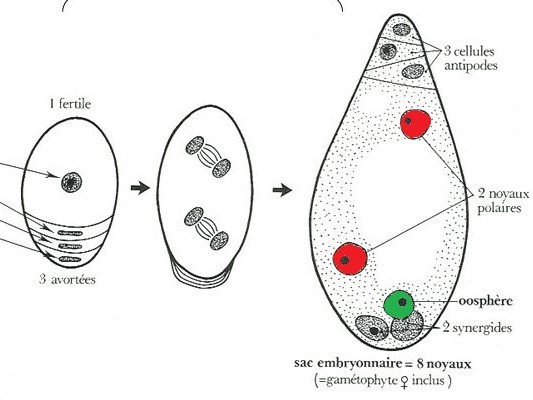
Figure 5.2: Développement du sac embryonnaire
Cette macrospore survivante se duplique ensuite de nombreuses fois pour former les 8 noyaux du sac embryonnaire. Les duplications (mitoses) sont des « copier-coller ». Ainsi, si par exemple le génome de la macrospore fertile est [jaune], alors l’oosphère, les noyaux polaires et toutes les autres cellules du sac embryonnaire porteront le même génome [jaune].
Ce processus de fabrication du sac embryonnaire a lieu pour chacun des grains qui composeront un épi de maïs. Ainsi, sur un même épi, environ la moitié des sacs embryonnaires porteront des cellules avec le génome [jaune] et l’autre moitié avec le génome [rouge].
5.2 Formation des gametes males
Dans notre exemple, la plante qui va produire les pollens porte dans son génome la paire de chromosomes n°1 sur laquelle on trouve le gène de la couleur de l’épi. Cette plante est ici hétérozygote sur le gène de la couleur, son génome est [jaune – rouge].
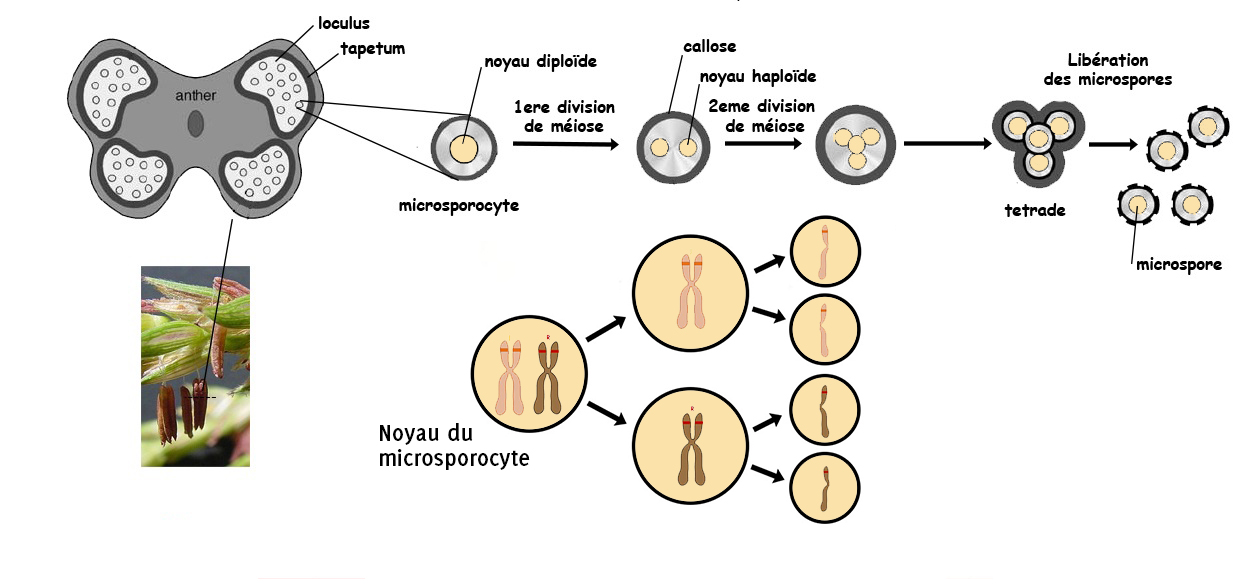
Le noyau du microsporocyte porte le même génome que la plante dans son noyau. Lors de la méiose, cette cellule va se diviser en deux, chacune des nouvelles cellules embarque un chromosome de la paire. Au moment de la seconde division cellulaire, les chromosomes sont coupés en deux, on obtient dont 4 cellules (les microspores) qui contiennent dans leur noyau une seule chromatide (un des deux bâtonnets qui composent le chromosome).
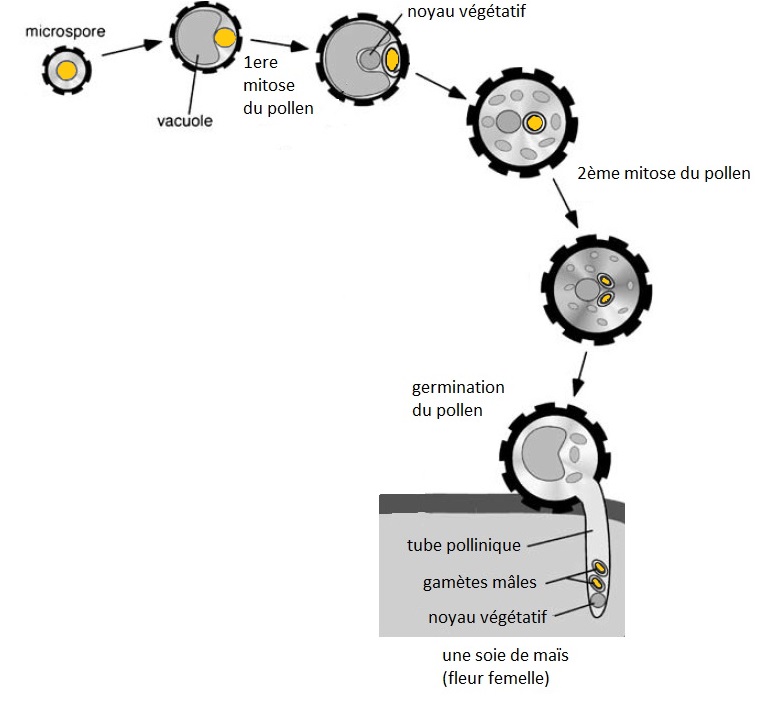
Lors de la maturation du grain de pollen, le noyau de la microspore est dupliqué pour former deux gamètes mâles parfaitement identiques.
Ainsi, dans le nuage de pollen que va produire cette plante, la moitié des grains de pollen porteront dans leurs 2 noyaux de gamètes le génome [jaune] et l’autre moitié le génome [rouge].
5.3 La fecondation
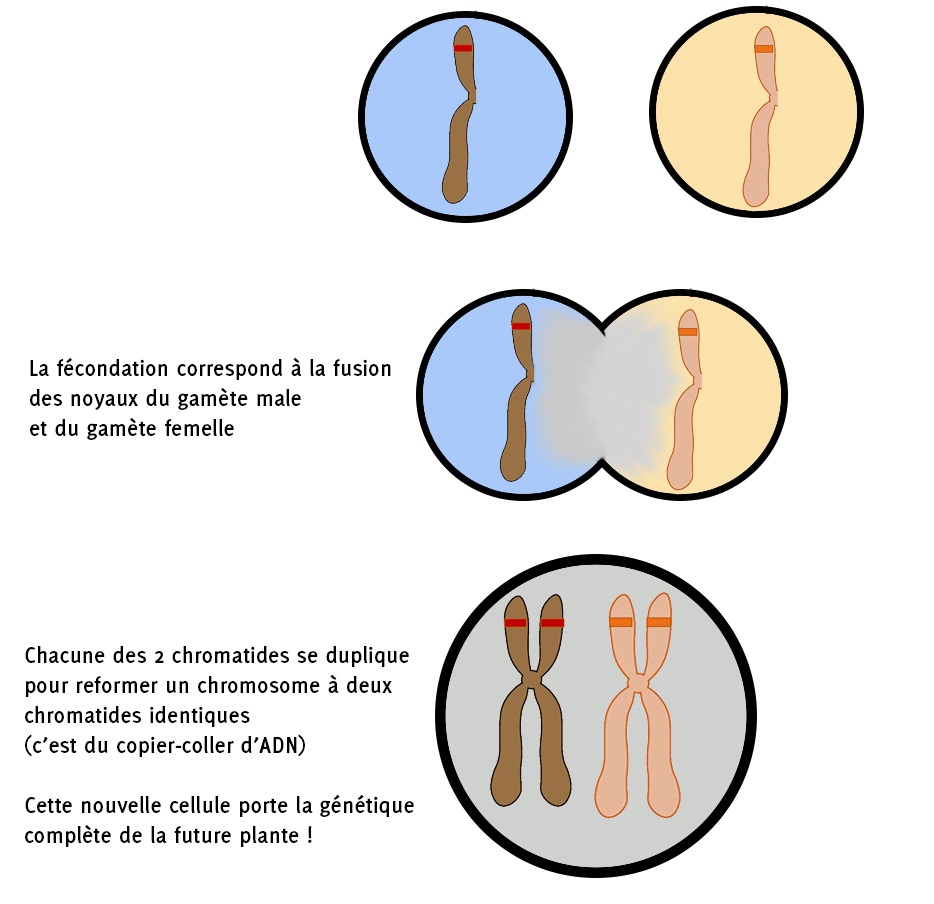
La fécondation est la rencontre et la fusion entre le gamète femelle et le gamète mâle (entouré en rouge ci-dessus). Cette nouvelle cellule est de nouveau diploïde : c’est la cellule embryonnaire qui va se diviser pour donner l’embryon de la graine, portant la génétique de la future plante. Ici nous avons illustré la fusion entre un gamète mâle portant le génome [jaune] et le gamète femelle portant le génome [rouge] ; le génome dans l’embryon et donc de la plante issue de ce grain sera hétérozygote [jaune-rouge].
Le deuxième gamète mâle fusionne avec les deux noyaux polaires du sac embryonnaire pour former la cellule souche à l’origine de l’endosperme, la chair du grain. La cellule souche de l’endosperme est donc triploïde.
Dans l’exemple où les cellules du sac embryonnaire sont issues d’une macrospore fertile de génétique [rouge], les noyaux polaires seront aussi de génétique [rouge] et donc, si le gamète mâle est de génétique [jaune], alors la fusion des deux noyaux polaires et du second gamète mâle donnera une cellule souche triploïde de génétique [rouge-rouge-jaune].
5.4 Hasard et probabilite lors d’un croisement
Si nous gardons l’exemple d’un individu dont je génome est hétérozygote [jaune-rouge] qui produit des pollens et vient féconder les soies d’un autre individu de même génome hétérozygote [jaune-rouge], on peut se poser la question suivante : quel sera le génome d’un grain de maïs quelconque issu de fécondation entre ces deux plantes ?
Il existe alors 4 scénarii différents, le génotype des descendants sera soit :
- Issu de la fécondation entre une oosphère portant l’information [jaune] et un pollen portant l’information [jaune]
- Issu de la fécondation entre une oosphère portant l’information [rouge] et un pollen portant l’information [jaune]
- Issu de la fécondation entre une oosphère portant l’information [rouge] et un pollen portant l’information [rouge]
- Issu de la fécondation entre une oosphère portant l’information [jaune] et un pollen portant l’information [rouge]
On peut résumer et calculer la probabilité d’obtenir chacun de ces génotypes grâce à un tableau de croisement.
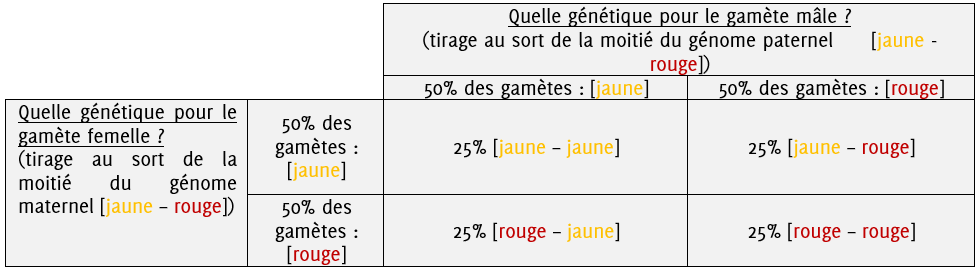
Dans cette situation, on a donc 50% de chance d’obtenir une plante à la génétique [rouge-jaune], 25% de chance d’obtenir une plante à la génétique [rouge-rouge] et 25% de chance d’obtenir une plante à la génétique [jaune-jaune].
5.5 Modele a 2 paires de chromosomes
L’étape clef commune à la formation des gamètes mâles ou des gamètes femelles est le moment où une cellule diploïde (donc avec les chromosomes représentés par paires) se divise par le processus de la méiose pour donner quatre cellules dont le génome est constitué d’une seule chromatide par type de chromosome. Nous avons observé le détail de cette transformation du génome lorsque celui-ci était réduit à une paire de chromosome. Regardons désormais ce qu’il se passe avec deux paires de chromosomes.
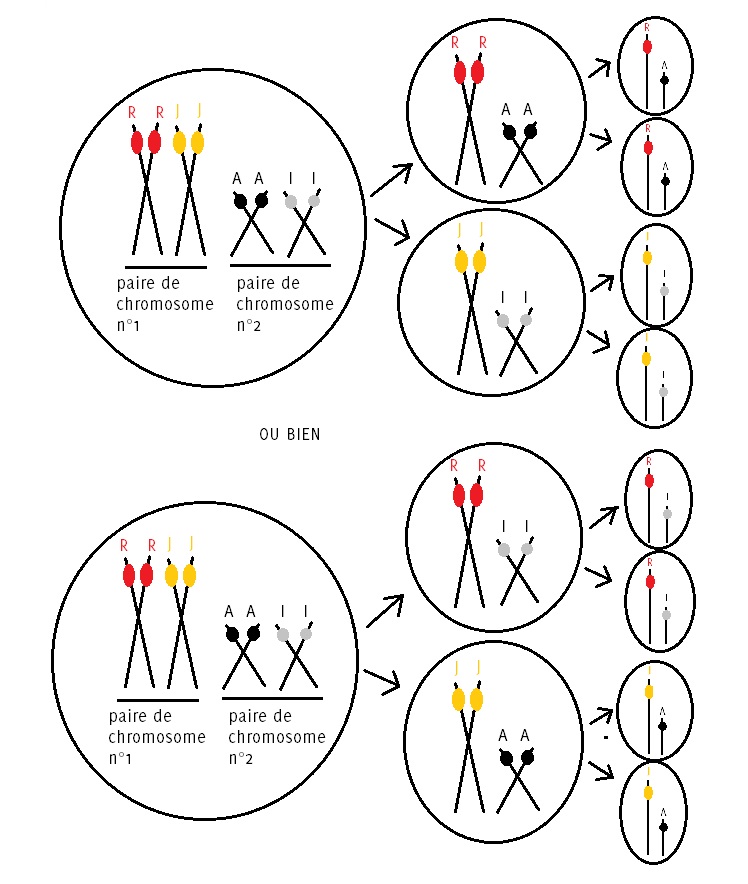
Nous introduisons dans notre exemple une nouvelle paire de chromosomes, la paire numéro 2. Nous représentons cette paire de chromosomes plus petite que la première pour bien la distinguer de celle-ci. Sur cette paire, nous imaginons qu’il existe un autre gène, appelons-le « régulateur », dont il existe deux versions différentes : l’allèle A et l’allèle I.
Un individu de maïs hétérozygote sur le gène de la couleur porté par la paire de chromosomes n°1 et hétérozygote sur le gène « régulateur » porté par la paire de chromosome n°2, donc de génome [jaune-rouge][A-I], commence une méiose à partir d’une cellule diploïde afin de produire des gamètes. Que ce soit des gamètes mâles ou des gamètes femelles, le raisonnement est le même.
Lors de la première division de méiose, les paires de chromosomes sont séparées dans deux cellules différentes. Il peut alors se passer deux choses : soit le chromosome [rouge] de la paire n°1 part en vadrouille avec le chromosome [A] de la paire n°2 et le chromosome [jaune] accompagne le chromosome [I] ou bien c’est l’inverse : le chromosome [rouge] s’en va avec le chromosome [I] et le chromosome [jaune] se coltine le chromosome [A].
Ainsi, cette plante peut produire 4 génotypes de gamète différents : [rouge][A], [rouge][I], [jaune][A] ou [jaune][I]. Si nous avions pris un modèle à 3 paires de chromosomes, une plante hétérozygote sur ces 3 paires pourrait produire alors 8 génotypes de gamète différents. Ainsi, avec un modèle à 10 paires de chromosomes hétérozygote, une plante pourrait produire 1024 génotypes de gamète différents.
Il existe d’autres subtilités génétiques, que nous ne présenterons pas ici, qui permettent d’augmenter la quantité de combinaisons génomiques lors de la formation des gamètes à tel point que les possibilités de combinaisons deviennent infinies et la probabilité d’obtenir deux gamètes au génome strictement identique est presque impossible.
La gamétogenèse est donc une formidable génératrice de diversité et la diversité des génomes issus de la fécondation des gamètes générés par deux individus différents (ou le même individu d’ailleurs !) est incommensurable. Ce bouillon de recombinaisons génétiques est nommé le « brassage génétique » et permet de créer des combinaisons génétiques sans cesse nouvelles dans une population d’individus diversifiés.
6 De la composition allelique au phenotype : l’expression des genes
L’expression des gènes, du génotype (l’information) au phénotype (la forme), se fait par des mécanismes physiologiques que nous avons décrits dans le premier chapitre « Rôle et fonctionnement biologique de l’expression des gènes ». Il n’est cependant pas forcément nécessaire de comprendre ces mécanismes dans le détail pour faire de la génétique appliquée (comme en sélection paysanne), seules quelques grandes formes d’interactions et d’expression des gènes sont à connaître.
Pour cela, nous allons reprendre notre exemple de maïs hétérozygote sur le gène « couleur de l’épi » qui possède des allèles différents : Jaune et Rouge.
Soit un des deux allèles est dominant, son expression prend donc le dessus sur l’autre allèle. L’allèle qui ne s’exprime pas est dit récessif.
Par exemple, si l’allèle [rouge] est dominant sur le [jaune], alors l’épi sera rouge. Si aucun des allèles n’est dominant, les deux s’expriment en même temps, on dit alors qu’ils sont additifs.
Par exemple, si l’allèle [rouge] et l’allèle [jaune] sont additifs, alors le phénotype sera le résultat de la somme des expressions des allèles : [jaune] + [rouge] = [orange]

Il existe aussi deux grands types de gènes :
Les gènes structurels : ils codent directement pour des caractères phénotypiques.
Les gènes de régulation : qui viennent activer ou désactiver l’expression des gènes structurels.
Pour illustrer le fonctionnement des gènes de régulation, nous allons prendre un nouvel exemple, toujours avec la couleur de l’épi.
On imagine le gène « couleur de l’épi » (présent sur la paire de chromosome n°1) avec les deux allèles suivants [jaune] et [rouge]. Les deux allèles sont additifs.
- Avant l’expression de ce gène, on considère que l’épi est blanc
- L’allèle [rouge] (R) permet la production du pigment rouge. -L’allèle [jaune] (J) permet la production du pigment jaune.
On imagine un nouveau gène (présent sur la paire de chromosome n°2), que nous allons appeler « inhibiteur de la couleur rouge de l’épi ». Ce gène possède deux allèles différents :
- L’allèle [actif] (A) qui vient inhiber la production du pigment rouge.
- Et l’allèle [inactif] (I) qui ne fait rien. L’allèle [actif] est dominant sur l’allèle [inactif].
Considérons une plante femelle (qui produit les soies) dont la génétique est [Rouge/Rouge] et [Actif/Inactif], pour simplifier nous allons écrire [RR] [AI]. Tentez de trouver la couleur de l’épi de cette plante.
Considérons une plante mâle (qui produit le pollen) dont la génétique est [Rouge/Jaune] et [Actif/Inactif], pour simplifier nous écrivons [RJ] [AI]. Tentez de trouver la couleur de l’épi de cette plante.
La réponse c’est…
- l’individu qui produit la fleur femelle présente un épi blanc : l’allèle [actif] du gène « Inhibiteur de la couleur rouge de l’épi » est présent et est dominant, son activité inhibe toute formation de pigment rouge, comme il n’y a pas d’allèle pour la production du pigment jaune, l’épi est blanc.
- l’individu qui produit le pollen présent un épi jaune : l’allèle [actif] du gène « Inhibiteur de la couleur rouge de l’épi » est présent et est dominant, son activité inhibe toute formation de pigment rouge mais pas celui du pigment jaune, l’épi est jaune.
Nous allons désormais dérouler un raisonnement pour prévoir le résultat du croisement entre ces deux plantes.
6.0.1 Formation des gamètes femelles
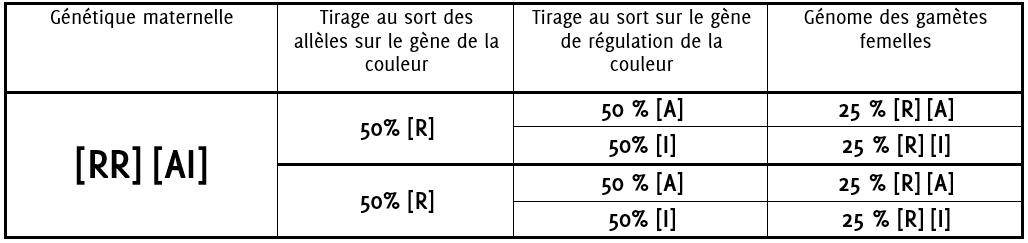
La plante femelle produira dans :
50% des cas des gamètes avec la génétique [R] [A] (25% + 25%)
50% des cas des gamètes avec la génétique [R] [I] (25% + 25%)
6.0.2 Formation des gamètes mâles
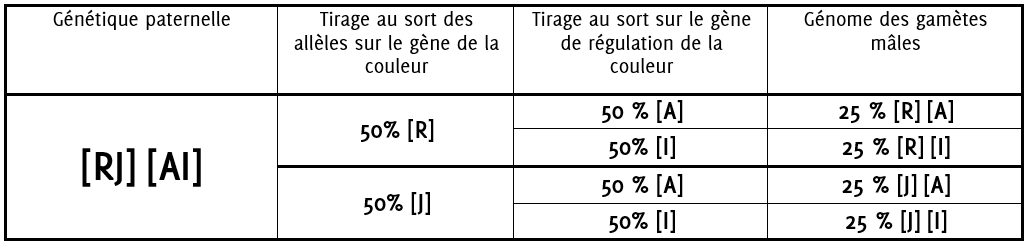
La plante mâle produira dans :
25% des cas des gamètes avec la génétique [R] [A]
25% des cas des gamètes avec la génétique [R] [I]
25% des cas des gamètes avec la génétique [J] [A]
25% des cas des gamètes avec la génétique [J] [I]
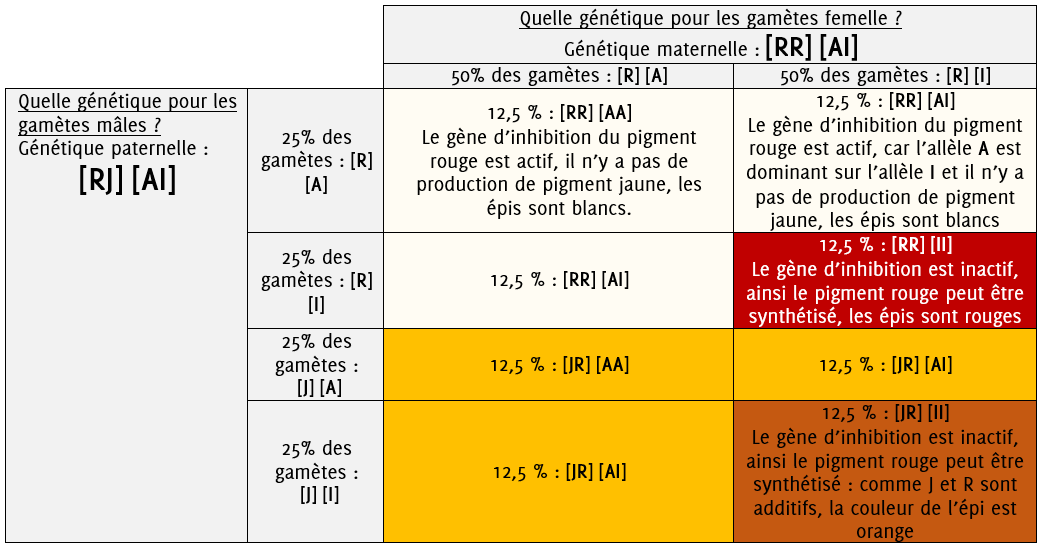
Figure 6.1: Fécondation
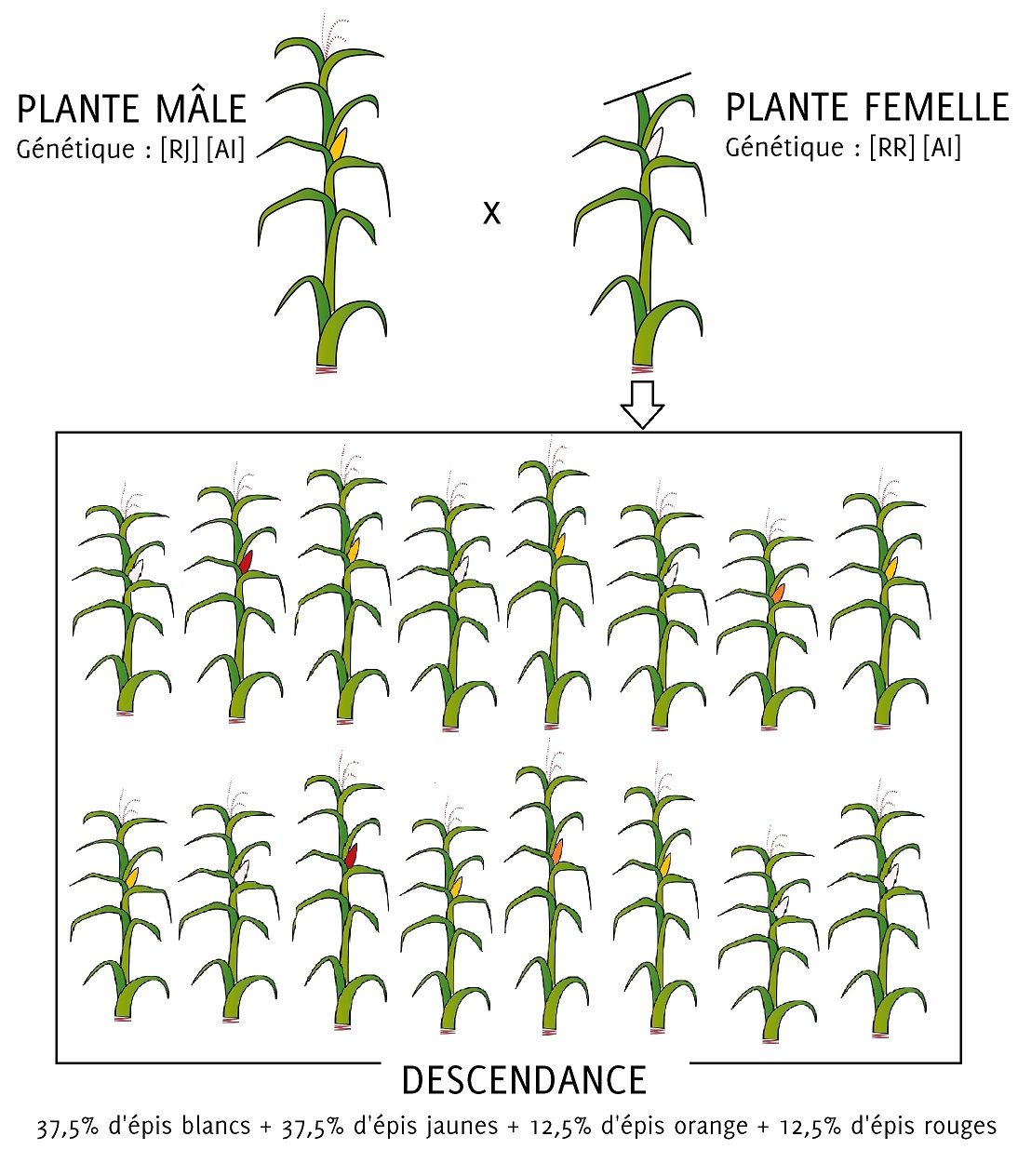
Figure 6.2: Voici ce qu’on observerait au champ si nous faisions ce croisement particulier : on voit, à travers cet exemple simple, que si on sème les graines d’un épi jaune, on peut tout de même trouver dans la descendance des épis rouges ou orange.
7 Conclusion
Par le système de production de gamètes et de fécondation, l’information génétique est transmise d’une génération à l’autre et permet ainsi de transmettre une partie des caractéristiques phénotypiques d’une population. Mais ce processus permet aussi de faire un nouveau brassage génétique (surtout chez les espèces allogames) qui va engendrer, par recombinaison aléatoire des allèles, des individus toujours uniques : ce brassage possède un pouvoir de création de diversité extrêmement puissant : un humain a par exemple la possibilité à lui seul d’engendrer plus 8 millions de gamètes différents !
Ainsi, la transmissibilité des caractères est régie par des phénomènes de hasard et de probabilité. Chercher à prédire le résultat d’une fécondation pour un individu en particulier au sein d’une population est hasardeux, cela revient à jouer au loto. Par contre, prédire le résultat d’un ensemble de fécondations sur une population de grande taille (plusieurs milliers d’individus) est parfaitement faisable : c’est comme si on effectuait tous les tirages possibles du loto en même temps : on n’a pas besoin de connaître le tirage pour savoir qu’on a gagné une fois! C’est avec ces principes que l’on peut prévoir une partie des résultats d’une sélection paysanne effectuée sur une variété diversifiée et allogame.
Enfin, grâce à la connaissance sur l’origine génétique des tissus du grain de maïs par le processus biologique de la double fécondation et à la connaissance sur les différentes formes d’expressions des allèles, il vous est désormais possible de comprendre une grande partie du codage génétique réel de la couleur des grains de maïs en lisant le chapitre bonus dédié : « Codage génétique de la couleur du grain de maïs ».
7.1 Ce qu’il faut retenir
La transmission des caractères d’une génération à une autre se fait principalement par la transmission de l’information génétique contenue dans le noyau de chaque cellule vivante. Cette transmission se fait en deux temps :
La formation des gamètes mâles et femelles. Les gamètes sont des cellules particulières car l’information génétique qu’elles contiennent est issue du tirage au sort de la moitié du génome de l’individu qui produit le gamète.
La fusion entre les gamètes mâle et femelle pour donner une cellule qui porte la génétique complète d’un nouvel individu : c’est la fécondation.
Le brassage génétique permet d’engendre des êtres vivants sans cesse différents
Un gène est une portion de l’ADN qui a un emplacement précis dans le génome (tel endroit de tel chromosome) et qui code pour un élément du phénotype (ex : couleur des yeux, largeur des feuilles, longueurs des poils,…). C’est presque toujours le concours de l’expression de nombreux gènes qui permet d’aboutir à un phénotype particulier. Il existe ainsi des gènes structurels et des gènes de régulation.
Les allèles sont des versions de gènes. Au sein d’une population, il peut exister une infinité de versions d’allèles pour un même gène mais un individu ne peut contenir dans son génome qu’un ou deux allèles différents : il est ainsi soit homozygote sur le gène, soit hétérozygote.
Lorsqu’un individu est hétérozygote sur un gène (2 allèles différents), les allèles peuvent être dominants, récessifs ou additifs.

Figure 7.1: Les incroyables aventures de la chromatide qui porte l’allèle R.
CHAPITRE SUIVANT : CHAPITRE 3 : TRANSMISSIBILITE DE L’INFORMATION GENETIQUE A L’ECHELLE D’UNE POPULATION ALLOGAME Ou l’évolution de la structure allélique
8 Source
[1] D. Twell, S.-A. Oh, et D. Honys, « Pollen Development, a Genetic and Transcriptomic View », in The Pollen Tube, vol. 3, R. Malhó, Éd. Berlin/Heidelberg: Springer-Verlag, 2006, p. 15‑45. doi: 10.1007/7089_042.
[2] P. A. Sabelli et B. A. Larkins, « The Development of Endosperm in Grasses », PLANT PHYSIOLOGY, vol. 149, no 1, p. 14‑26, janv. 2009, doi: 10.1104/pp.108.129437.
9 Document realise dans le cadre du projet COVALIENCE
NOEL Robin (2021). Chapitre 2 : La sexualité du maïs, la transmission des gènes d’une génération à l’autre et le brassage génétique. Projet Casdar Covalience. 15 pages.
Contact mail auteur : biodiversite@agrobioperigord.fr