- 1 CHAPITRE 4 : LES FORCES QUI FONT EVOLUER LES STRUCTURES ALLELIQUES - Mutation, migration, derive genetique, selection naturelle et consciente
- 2 Les forces qui engendrent de la diversite allelique au sein d’une population
- 3 Les forces qui reduisent de la diversite allelique au sein d’une population
- 4 Equilibre dans la structure allelique d’une population
- 5 Ce qu’il faut retenir
- 6 Sources
- 7 Document realise dans le cadre du projet COVALIENCE
CHAPITRE PRECEDENT : CHAPITRE 3 : TRANSMISSIBILITE DE L’INFORMATION GENETIQUE A L’ECHELLE D’UNE POPULATION ALLOGAME Ou l’évolution de la structure allélique
1 CHAPITRE 4 : LES FORCES QUI FONT EVOLUER LES STRUCTURES ALLELIQUES - Mutation, migration, derive genetique, selection naturelle et consciente
1.1 Preambule
Certaines notions définies dans les chapitres précédents sont utiles pour comprendre le chapitre 4. Nous avons vu dans les chapitres précédents que le phénotype des individus était en grande partie dû à la nature et à la structure des gènes stockés sous la forme d’ADN dans le noyau des cellules des êtres vivants et concaténés en chromosomes. Il peut exister plusieurs versions différentes pour un gène, on nomme ces versions de gène les allèles. Grâce à la formation des gamètes et à la fécondation, la génétique se transmet d’une génération à l’autre et les allèles sont sans cesse recombinés à l’échelle d’une population : c’est le brassage génétique.
A l’échelle d’une population, on peut prédire et décrire les proportions des différentes combinaisons alléliques, et donc la proportion des phénotypes, grâce aux équations de Hardy-Weinberg. La diversité des allèles et leur proportion relative dans une population est nommée « structure allélique ». La structure allélique est un synonyme de diversité génétique.
1.2 Pourquoi lire ce chapitre ?
Dans ce chapitre nous allons présenter les grandes forces naturelles (ou non) qui font évoluer dans le temps la structure allélique d’une population de plante cultivée allogame comme le maïs. Certaines de ces forces font augmenter la diversité génétique d’une population et d’autres forces la font diminuer. Connaître et comprendre la nature de ces mécanismes permet au paysan sélectionneur d’adopter des raisonnements qui peuvent aboutir à des techniques particulières de sélection et multiplication du maïs en fonction des objectifs et affinités de celui-ci.
Bien au-delà de la sélection paysanne, ce chapitre et les suivants permettront d’introduire des notions structurantes pour les disciplines qui touchent à la diversité génétique et à l’évolution du vivant : sauvegarde de la biodiversité cultivée, biologie de conservation des espèces naturelles, sélection naturelle et évolution des espèces,…
2 Les forces qui engendrent de la diversite allelique au sein d’une population
Un gène est une séquence d’ADN définie arbitrairement qui occupe une place précise sur une paire de chromosomes du génotype. Comme nous l’avons vu dans le premier chapitre, on associe souvent un gène à la fabrication d’une protéine donnée qui jouera un rôle précis dans l’organisme et le phénotype résulte donc du concours des actions des protéines. Mais pour un gène donné, il peut exister différentes séquences d’ADN qui engendreront des protéines légèrement différentes avec des actions métaboliques différentes. Ces différentes séquences sont nommées les allèles. Il peut exister une grande quantité d’allèles différents pour un même gène dans une population mais nous rappelons qu’un individu au génome diploïde ne peut posséder qu’un à deux allèles différents par gène.
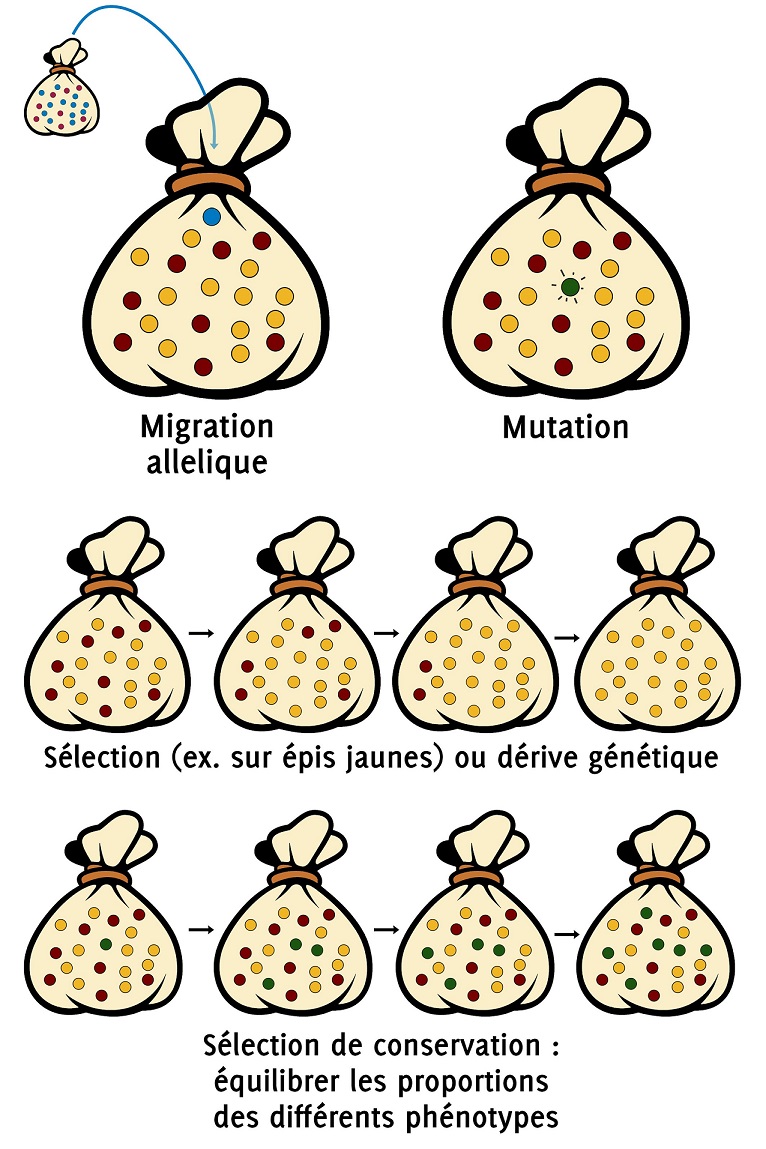
La diversité génétique d’une population peut donc être décrite grâce à la diversité des allèles que l’on trouve dans cette population. L’augmentation de la diversité allélique nécessite donc l’apparition de nouveaux allèles dans une population. Il existe deux mécanismes qui permettent l’enrichissement allélique dans une population : les migrations et les mutations.
2.1 La migration allelique
Comme son nom l’indique, la migration allélique décrit un enrichissement de la diversité génétique par un apport extérieur à la population.
Pour une population de maïs par exemple, cet apport peut-être le résultat de la fécondation des soies par des pollens issus d’une autre population et apportés par le vent. Il peut aussi venir d’un mélange de graines dans les sacs de semences avant le semis.
Selon les objectifs du paysan multiplicateur, la migration allélique peut être perçue positivement ; il existe par exemple une pratique paysanne traditionnelle qui consiste à réintégrer, en petite quantité et de temps en temps, de la génétique « étrangère » dans une population.

Parfois, cette migration allélique peut-être perçue négativement car elle apporte des caractéristiques phénotypiques non désirées par le paysan et on en parle alors comme d’une « pollution » ou d’une « contamination ». C’est le cas par exemple lorsque des pollens de maïs OGM étasuniens fécondèrent des populations traditionnelles de maïs mexicains dans les années 2000 [1] ou plus communément lorsqu’une population aux grains blancs présente quelques grains jaunes issus d’une pollinisation par une parcelle voisine de maïs au grain jaune (comme sur la photo ci-dessus avec une population de maïs blanc « Grand Cachalut » semée trop proche d’une variété de maïs au grain jaune).
Dans la plupart des situations, les migrations alléliques n’engendrent que des modifications minimes dans la structure allélique de la population car les nouveaux allèles ne seront représentés qu’en toutes petites proportions par rapport aux allèles déjà existants dans la population. Cependant certaines forces évolutives comme le hasard ou la sélection, que nous décrirons en détail plus tard, peuvent permettre la modification de la proportion des allèles nouvellement arrivés.
2.2 Les mutations
Dans la vie d’un animal, d’une plante ou d’un champignon, l’organisme se développe et assure ses fonctions vitales grâce à la division cellulaire. Chaque jour notre corps, ou un plant de maïs, fabrique de nouvelles cellules à partir des cellules existantes. L’ADN, présent dans le noyau de chacune des cellules, doit alors se répliquer lors de cette division, faire un « copier-coller ». Plusieurs protéines spécialisées sont chargées de réaliser cette réplication, c’est comme une petite équipe de moines copistes qui œuvre d’arrache-pied pour conserver le sacro-saint code ADN de l’organisme des affres du temps.

Mais parfois, et cela arrive assez souvent, les protéines copistes ont un peu mal aux yeux et se trompent dans le recopiage de la séquence d’ADN. Cela peut être une modification, un oubli ou un ajout de base azotée (rappelons que les bases azotées : G, C, A, T composent l’ADN). Ainsi une séquence d’ADN qui ferait GTT CAG TAC TGA pourrait devenir GTA CAG TAC TGA.
Comme un allèle est par définition une séquence d’ADN spécifique, une modification dans la séquence engendre un nouvel allèle. C’est ce qu’on appelle une mutation. Cette mutation peut par exemple intervenir au moment des divisions cellulaires qui engendre les gamètes (Cf. chapitre 2). Si c’est le cas, un allèle muté sera alors transmis à la descendance et une nouvelle diversité génétique apparaîtra dans la population.
Les mutations sont parfaitement aléatoires, les organismes vivants n’engendrent pas des mutations génétiques par dessein, c’est un mécanisme aveugle qui peut toucher absolument tous les gènes. Mais la fréquence des mutations peut dépendre de l’environnement comme par exemple l’exposition à de la radioactivité.
2.3 Consequences des mutations sur le phenotype
Certaines mutations sont dites silencieuses car même s’il y a une petite différence dans la séquence d’ADN, il n’y a pas de modification dans la séquence des acides aminés qui compose les protéines (Cf. chapitre 1).
D’autres fois, la mutation engendre une légère modification de la protéine codée mais cette modification ne perturbe pas le fonctionnement de la protéine, le phénotype n’est donc pas affecté.
Et enfin, il arrive que la mutation engendre une véritable altération de forme de protéine dont le fonctionnement sera alors modifié, ce qui aura une conséquence sur le phénotype. Ces conséquences peuvent prendre une grande diversité de formes et d’intensités : elles peuvent parfois apporter un petit avantage reproductif dans certaines conditions de température ou d’humidité, peuvent engendrer une modification de la nature d’un pigment de couleur, peuvent modifier les régulations hormonales de la plante et provoquer du nanisme ou du gigantisme ou, parfois même, perturbent tellement le fonctionnement de l’organisme qu’elles entraînent la mort prématurée de l’individu. Dans ce dernier cas on dit que le gène muté est délétère.

Exemple ci-dessus (à gauche) des effets d’une mutation génétique perturbant la synthèse de la chlorophylle sur un pied de maïs population observé en Dordogne en 2020. Certaines mutations peuvent engendrer des pieds complètement albinos, des épis avec plusieurs pointes, des épis multiples (exemple à droite dans une population de la variété Bagan) des anomalies florales (grains sur la panicules ou fleur mâle au niveau de l’épi) comme sur la photo ci-dessous. Nota : de nombreuses variétés de plantes ornementales sont par exemples issues d’un isolement de jolies plantes mutées (souvent d’ailleurs sur l’albinisme partiel des feuilles).

Comme pour la migration allélique, les mutations n’engendrent que des modifications minimes dans la structure allélique de la population car les nouveaux allèles ne seront représentés qu’en toutes petites proportions par rapport aux allèles déjà existants dans la population. De plus les allèles mutés qui engendrent des conséquences délétères ne seront pas transmis aux générations suivantes (car l’individu meure avant de produire des pollens ou grains). Dans l’histoire de la domestication des plantes, les paysans ont su repérer dans le peuplement végétal, isoler et reproduire des plantes « intéressantes » issues de mutations. C’est ainsi qu’une formidable diversité de variétés fut engendrée par accumulation successive de petites mutations grâce à la multiplication récurrente des plantes, la sélection paysanne et la pluralité des communautés paysannes.
3 Les forces qui reduisent de la diversite allelique au sein d’une population
Les proportions relatives des allèles pour un gène donné dans une population ne sont pas stables : il existe des forces qui viennent modifier la fréquence des allèles, donc la structure allélique de la population, au fil des générations. Ainsi, aux marges de ces modifications et fluctuations de proportions d’allèles, il est possible que des allèles viennent à disparaître complètement de la population, réduisant ainsi la diversité allélique de la population.
Il existe deux grandes forces qui agissent en permanence sur la structure allélique des populations : il s’agit de la sélection (naturelle ou par la main de l’humain) et, plus étonnamment, de l’unique effet du hasard que l’on appelle « dérive génétique ».
3.1 La selection
Le terme « sélection » induit l’idée d’un choix, d’une ségrégation, d’un casting du vivant, il y a ceux qui seront « sélectionnés » et ceux qui ne le seront pas. Cette sélection est opérée par l’environnement sur les phénotypes.
Par exemple, dans un environnement agricole où circule un ravageur des cultures, les individus avec un phénotype « résistant » auront plus de chance d’attendre l’âge de reproduction que les individus avec un phénotype « non résistant ». Si les phénotypes « résistants » et « non résistants » sont induits par des allèles spécifiques, alors les allèles jouant un rôle dans la résistance auront plus de chance d’être transmis à la génération suivante que les allèles ne conférant pas cette résistance. Ainsi, dans cette situation, l’environnement exerce une pression de sélection qui favorise les phénotypes « résistants » et défavorise les phénotypes « non résistants » : au fil des générations, la proportion des allèles associés au phénotype « résistant » augmentera et la proportion des allèles associés au phénotype « non résistant » diminuera jusqu’à, possiblement complètement disparaître de la population.
La main du sélectionneur est une pression de sélection supplémentaire qui induit aussi des modifications dans les proportions alléliques et qui peut, de même, aboutir à la disparition d’allèles non désirés (ex : l’allèle induisant la couleur jaune du grain de maïs si le sélectionneur souhaite avoir une population entièrement blanche).
Les chapitres qui suivront permettront d’aller plus loin sur les notions de sélection naturelle des plantes cultivée et de sélection paysanne.
3.2 Le hasard et la derive genetique
Dans le chapitre 3, nous avons vu que lors de la formation des gamètes mâles et femelles, le génome se divise en deux pour former des cellules sexuelles haploïdes (un seul chromosome par type de chromosome au lieu de deux). Dans ce processus de formation des gamètes et de fécondation, il est possible que certains allèles ne soient pas transmis à la génération suivante par le simple fait du hasard.
Pour le comprendre simplement, prenons une toute petite population d’humain fermée (aucune entrée possible de nouvelle personne) constituée de deux individus dont :
un homme dont le génotype sur le chromosome 21 est [X – Y]
une femme dont le génotype sur le chromosome 21 est [X – X]
Ces deux individus engendrent, par exemple, 5 descendants, tous issus du processus de formation des gamètes et de fécondation.
Il existe une probabilité pour que ces 5 descendants n’aient hérités de leurs parents que des chromosomes X, chacun a possède donc un génotype [X – X]. Les 5 descendants ici sont alors des filles. Sur cette toute petite population, l’allèle Y a disparu en une génération sans aucune forme de pression de l’environnement.
Ce simple exemple nous permet d’illustrer le fait que le brassage génétique permis par la reproduction sexuelle (formations des gamètes et fécondations) induit, à l’échelle des populations, des modifications de la structure allélique sans qu’il y ait besoin de pression de l’environnement.
Pour une population fermée (sans migration allélique) et sans sélection (naturelle ou consciente), le brassage allélique induit sur des temps très longs la disparition d’allèles, et donc l’homozygotie (2 même versions d’allèle sur les deux chromosomes formant une paire), sur absolument tous les gènes. Cette tendance est absolument inéluctable : c’est la dérive génétique.
Certaines conditions peuvent accélérer la dérive génétique comme par exemple le taux d’autofécondation (plus il est fort, plus la dérive est rapide) ou la taille de la population (plus elle est grande plus la dérive est lente).
Un chapitre complet sera consacré à la dérive génétique sur les espèces cultivées allogames comme le maïs.
4 Equilibre dans la structure allelique d’une population
La diversité allélique d’une population est le résultat d’un équilibre mouvant entre l’apparition de nouveaux allèles par migration ou mutation et la disparition d’allèles par dérive génétique ou sélection. Ces quatre grandes forces régissent l’évolution du vivant, engendrent l’apparition et la disparition d’espèces depuis l’apparition de l’ADN sur terre. La sélection paysanne et la domestication des plantes et des animaux permettent d’accentuer ces forces et d’engendrer plus rapidement de nouvelles diversités pour adapter le vivant à la diversité des besoins, des envies et des sensibilités de l’espèce humaine.

Dans certaines situations particulières, on peut parfois considérer que la sélection paysanne permet d’augmenter la diversité génétique d’une population mais sans création de nouvelle diversité allélique.
La diversité d’un système (que ce soit la structure allélique d’une population, un écosystème, une caisse de jouets, une remorque de gravier) peut être évaluée en décrivant le nombre d’éléments différents qu’on peut y trouver (10 000 allèles différentes, 20 espèces d’oiseaux, 12 jouets différents, 3 types de roches) mais aussi en décrivant la proportion relative des différents éléments du système. Pour mieux comprendre ce point, nous pouvons imaginer deux écosystèmes marins différents dont nous souhaitons évaluer l’état de la biodiversité.

Le premier écosystème est composé de 990 sardines et de 10 maquereaux. Le second écosystème est composé de 500 sardines et 500 maquereaux.
Même si dans les deux situations, on compte une richesse spécifique de 2 espèces (sardine et maquereau), l’état de la biodiversité du second écosystème semble « meilleur » que celui du premier car les effectifs entre les espèces sont plus équilibrés. Il existe différents indicateurs mathématiques qui permettent ainsi d’évaluer plus finement la diversité d’un écosystème au-delà du nombre d’espèces qui le compose ou la diversité allélique d’une population au-delà du nombre d’allèles différents.
Ainsi, la sélection peut permettre de rééquilibrer des proportions relatives de phénotypes dans une population et d’augmenter la variation génétique de cette population en privilégiant les phénotypes rares pour constituer la génération suivante.
4.1 Pour aller plus loin
L’index de Nei (noté HS) est souvent le plus utilisé et le plus facile à calculer et interpréter pour étudier la variation génétique (ou autres types de variations) d’une population. En voici la formule où i un élément particulier et pi la proportion de cet élément. HS correspond à 1 moins la somme des proportions des éléments au carré.
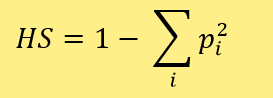
Dans l’exemple des écosystèmes marins : l’index de Nei pour le premier écosystème HS = 1 – ((0.990)2 + (0.01)2) = 0.0198 et pour le second écosystème HS = 1 – ((0.5)2 +(0.5)2) = 0.5
5 Ce qu’il faut retenir
La structure allélique d’une population permet de définir plus précisément ce qu’est la diversité génétique.
Cette structure allélique se définit par la diversité des allèles présents dans une population et la proportion relative de ces allèles. Mais la structure allélique n’est pas figée : elle évolue sous l’effet de 4 grandes forces.
Les forces qui augmentent la diversité allélique sont :
la migration allélique (qui peut être perçue positivement ou négativement selon les contextes)
les mutations génétiques qui sont l’unique forme de création de nouveaux allèles. Ces mutations sont aléatoires et peuvent avoir des effets nuls, positifs ou négatifs.
Les forces qui peuvent entraîner la perte d’allèles dans une population sont :
la dérive génétique qui est une force inéluctable liée au hasard produit par la formation des gamètes et le brassage génétique
la sélection (naturelle ou consciente) qui peut par exemple permettre de supprimer dans une population des allèles indésirables
La sélection peut cependant permettre de contrecarrer les effets de la sélection naturelle ou de la dérive génétique en privilégiant les phénotypes rares pour constituer la génération suivante. Ces 4 forces agissent de concert et constituent un équilibre dynamique. De manière générale, une perte d’allèles trop rapide et trop récurrente engendre de la consanguinité qui peut mener, dans certains cas extrêmes, à la disparition d’une population.
CHAPITRE SUIVANT : CHAPITRE 5 : L’UNIQUE EFFET DU HASARD SUR LA STRUCTURE ALLELIQUE - Derive genetique, etranglement, consanguinite et nombre minimum d’individus a selectionner
6 Sources
[1] J. Stolz, « Au Mexique, le berceau du maïs contaminé par des OGM », Le Monde, déc. 11, 2008. [En ligne]. Disponible sur: https://www.lemonde.fr/planete/article/2008/12/11/au-mexique-le-berceau-du-mais-contamine-par-des-ogm_1129750_3244.html
7 Document realise dans le cadre du projet COVALIENCE
NOEL Robin (2021). Chapitre 4 : les forces qui font évoluer les structures alléliques. Mutation, migration, dérive génétique, sélection naturelle et consciente. Projet Casdar Covalience. 7 pages.
Contact mail auteur : biodiversite@agrobioperigord.fr