Cet article fait appel à des notions de génétique de base comme “gènes”,“allèles”,“mutation”,“brassage génétique”,“phénotype”…la lectures des articles sur les bases de la génétique “Du génotype au phénotype” et “Organisation, fonctionnement et transmissibilité des gênes” sont conseillés.

La sélection paysanne est une sélection effectuée par et pour les agriculteurs. Elle est l’effet combiné de deux processus de modification de la structure génétique des populations de plantes cultivées : la sélection naturelle (on parle aussi souvent d’adaptation, ex : à un terroir) et la sélection consciente. Dans cet article, nous développerons la notion de sélection naturelle.
Evolution ou adaptation
L’évolution ou l’adaptation d’une population c’est quand la génétique de cette population évolue au cours du temps sous l’effet combiné du hasard (brassage génétique et mutation), de flux de gênes (croisement, contaminations…) et de la pression de l’environnement (la sélection naturelle).
Comprendre le principe de l’evolution avec un papillon
La Phalène du bouleau est un papillon de nuit européen qui constitue un très bon exemple pour expliquer les grands principes de la sélection naturelle et illustrer l’apparition d’une innovation morphologique avantageuse due à une mutation génétique.

Figure 1: La phalène du bouleau (Biston betularia) habitus sauvage
Dans les années 1830, la Grande Bretagne s’industrialise à grande vitesse, c’est l’ouverture des grandes mines à charbon, du travail à la chaine, des industries, des usines et des révolutions technologiques. Cette effervescence de production entraîne, notamment à cause du charbon, carburant essentiel des moteurs d’usines, un important dégagement de suie dans l’environnement, noircissant le paysage.
Or, la Phalène du bouleau joue sur sa couleur et ses dessins pour se camoufler le jour en se posant sur les troncs de bouleaux. Les bouleaux devenus noirs, les phalènes deviennent alors la cible d’une importante prédation par les oiseaux car leur mimétisme n’est plus efficace.
Dans la population de phalènes, lors d’une reproduction, un gène mute chez un individu. Une mutation c’est comme une erreur de traduction dans la transmission de l’information génétique d’une génération à une autre (cette mutation ayant très probablement déjà eut lieu auparavant et à plusieurs reprises mais est restée inaperçue à cause de son “inutilité” évolutive). L’allèle muté du gêne est dominant sur l’allèle sauvage initial et entraîne une importante modification phénotypique: à savoir le changement de coloration du corps et des ailes du blanc au noir.
(Attention : l’environnement n’influe ni sur le nombre, ni le type de mutation, excepté la radioactivité, ces mutations interviennent absolument aléatoirement).
La nouvelle forme de phalène du bouleau, f.carbonaria, présente une livrée (aspect visuel présenté par un insecte) bien mieux adaptée à cet environnement noirci et devient donc beaucoup moins exposée aux prédateurs. La sélection naturelle opérant, le rapport f.carbonaria / habitus typique passe de 0/100 en 1860 à 96/4 en 1948. La population de phalènes est donc devenue très majoritairement noire.
En 1950, la Grande Bretagne met en œuvre un programme de dépollution réduisant notamment les émissions de suie, les bouleaux redeviennent blancs et la forme noire de la phalène commence à avoir du mal à se cacher… En 50ans, le pourcentage de la population de f.corbonaria est passé de 95% à 15%, montrant bien le poids et l’influence de la sélection naturelle sur une population donnée et les interactions génétique/environnement sur la capacité de certains individus à se reproduire plus facilement que leurs congénères.
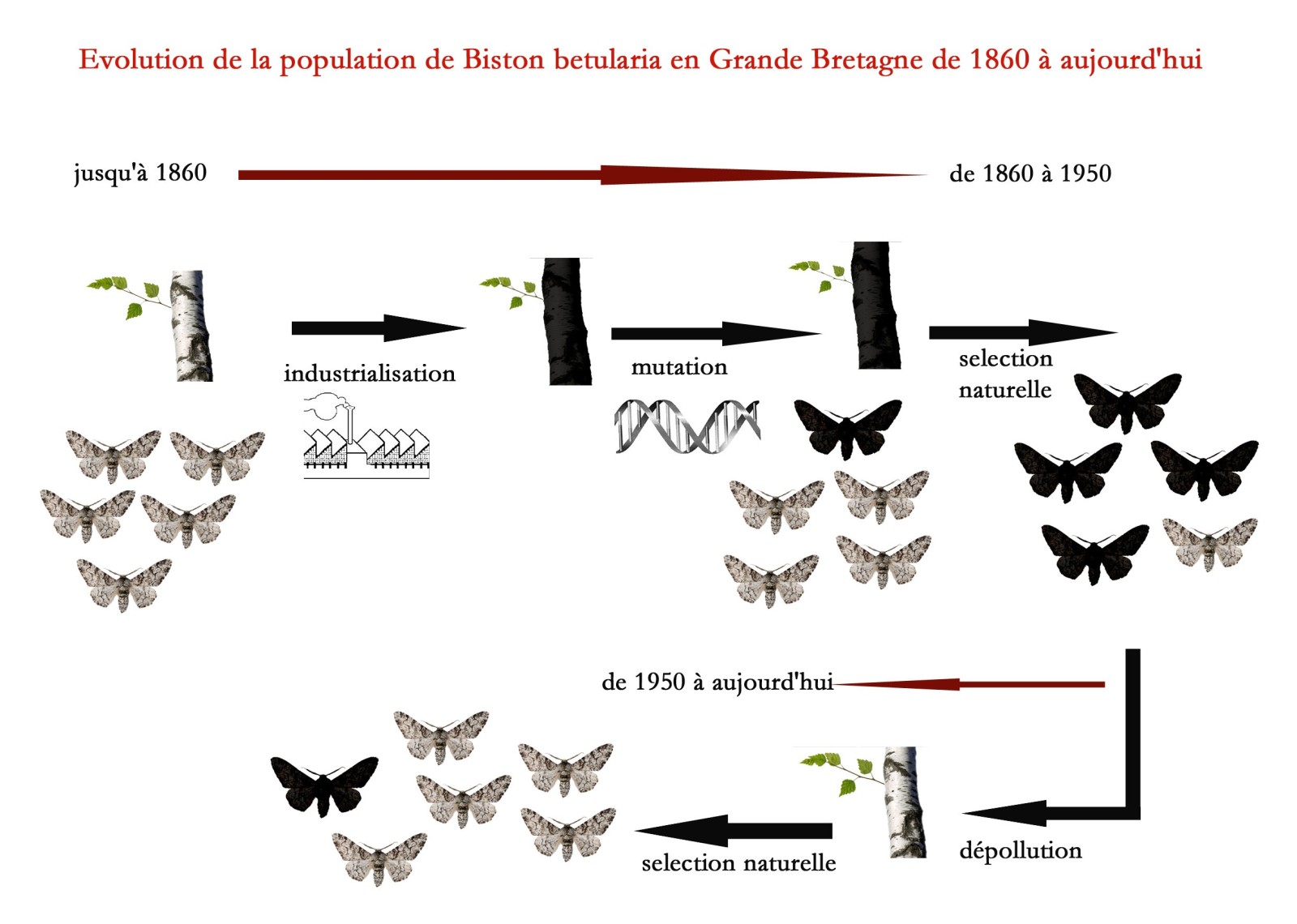
Figure 2: Le processus de sélection naturelle de la phalène du Bouleau en GB
L’adaptation c’est l’augmentation de la capacite d’une plante a produire une descendance viable
La sélection naturelle, telle qu’elle est décrite par Darwin, peut se définir comme un avantage reproductif procuré par les conditions de l’environnement aux individus ayant un caractère avantageux vis-à-vis de cet environnement et leur assurant une descendance plus importante que les individus n’ayant pas ce caractère (l’exemple du papillon).
Dans le cadre des productions végétales, on peut définir le concept de sélection naturelle en un processus qui favorise la transmission d’allèles (versions de gènes : voir article “Organisation, fonctionnement et transmissibilité des gênes”) d’une génération à une autre, lorsque le génome (ensemble de l’information génétique) des individus, dans un environnement donné, augmente:
- la capacité à germer
- la probabilité de survie jusqu’à l’âge de reproduction
- le nombre de graines produites [1]
Comme l’environnement agricole dans lequel évoluent les espèces cultivées est un milieu anthropique, le terme de « sélection naturelle » peut être discuté bien qu’il soit communément utilisé dans la littérature scientifique et la littérature grise. Par ce concept, il faut entendre « sélection du/par le milieu » ou « sélection de/par l’environnement ». Les plantes cultivées évoluent dans un agroécosystème constitué « de différents éléments tels que plantes, animaux, sol, climat et leurs interactions, en intégrant aussi l’effet du management agricole humain sur ces éléments »[2]. Conformément à la définition du système de culture de Sebillotte (1974)[3], le management agricole humain peut être décomposé en trois éléments : la nature des cultures, l’ordre de succession des cultures et les itinéraires techniques appliqués à ces différentes cultures.
La sélection naturelle est donc inscrite dans des interactions complexes entre génétique et environnement (figure 3). De plus, chaque individu est inclu dans un peuplement végétal : les plantes sont en interaction les unes avec les autres (compétition pour la lumière, nutriments, …) et l’intensité de ces interactions dépend d’éléments de l’agroécosystème (fertilisation, densité de semis…).
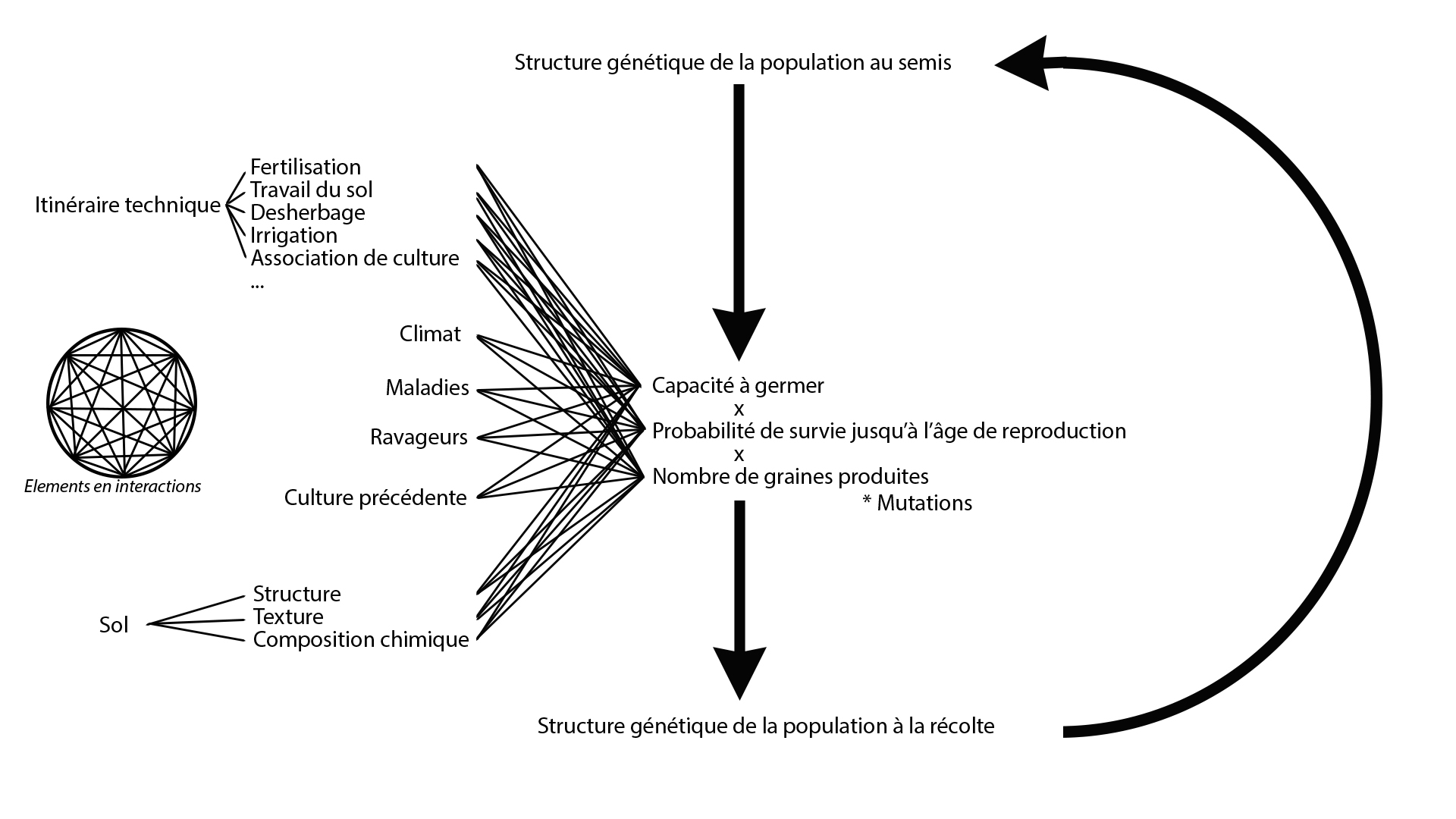
Figure 3: La sélection naturelle sur les plantes cultivées
L’application de la théorie de la sélection naturelle à la multiplication récurrente de semences dans un agroécosystème donné permet d’affirmer que : comme la sélection naturelle favorise les individus capables de produire le plus de descendance viable (nombre de grains), l’évolution génétique des populations tend alors à favoriser les individus les plus productifs sur la composante de rendement nombre de grains par pied et donc potentiellement le rendement grain de la culture.
Etat adapte et processus d’adaptation
David (1992) a observé l’adaptation d’une population de blé à un environnement donné : cette adaptation est caractérisée ici par une augmentation du rendement et une modification de la structure allélique de la population [4], soit une évolution dans les proportions relatives des allèles dans la population d’une génération à une autre. D’autres auteurs montrent des évolutions dans les structures alléliques des populations en lien avec l’apparition de résistance de l’orge au mildiou sur 9 cycles de sélection [5] ou du blé sur 8 cycles de sélection [6].
L’adaptation d’une population à un agroécosystème dépend, entre autre, de la pression de l’environnement sur la population. Hartwig et al. (1982) ont montré qu’une population de soja cultivée dans un environnement infesté de nématodes est passée d’une proportion initiale de 5% de plantes résistantes en génération 1 à 40% en génération 5, tandis que la proportion de plantes résistantes n’a pas évoluée sur le témoin cultivé dans un environnement sain[7].
L’adaptation des populations à un environnement est le plus souvent étudiée sans l’usage de marqueurs moléculaires et donc sans suivi de l’évolution de la structure allélique des populations. Ces études passent l’analyse des interactions cultivar x environnement pour différents cultivars issus d’une même population et sélectionnés/reproduits dans des environnements distincts (figure: 4) ou plus simplement par la comparaison de cultivars dans des environnements différents (étude de l’état « adapté » et non du processus d’adaptation car on considère qu’il existe toujours un ancêtre commun).
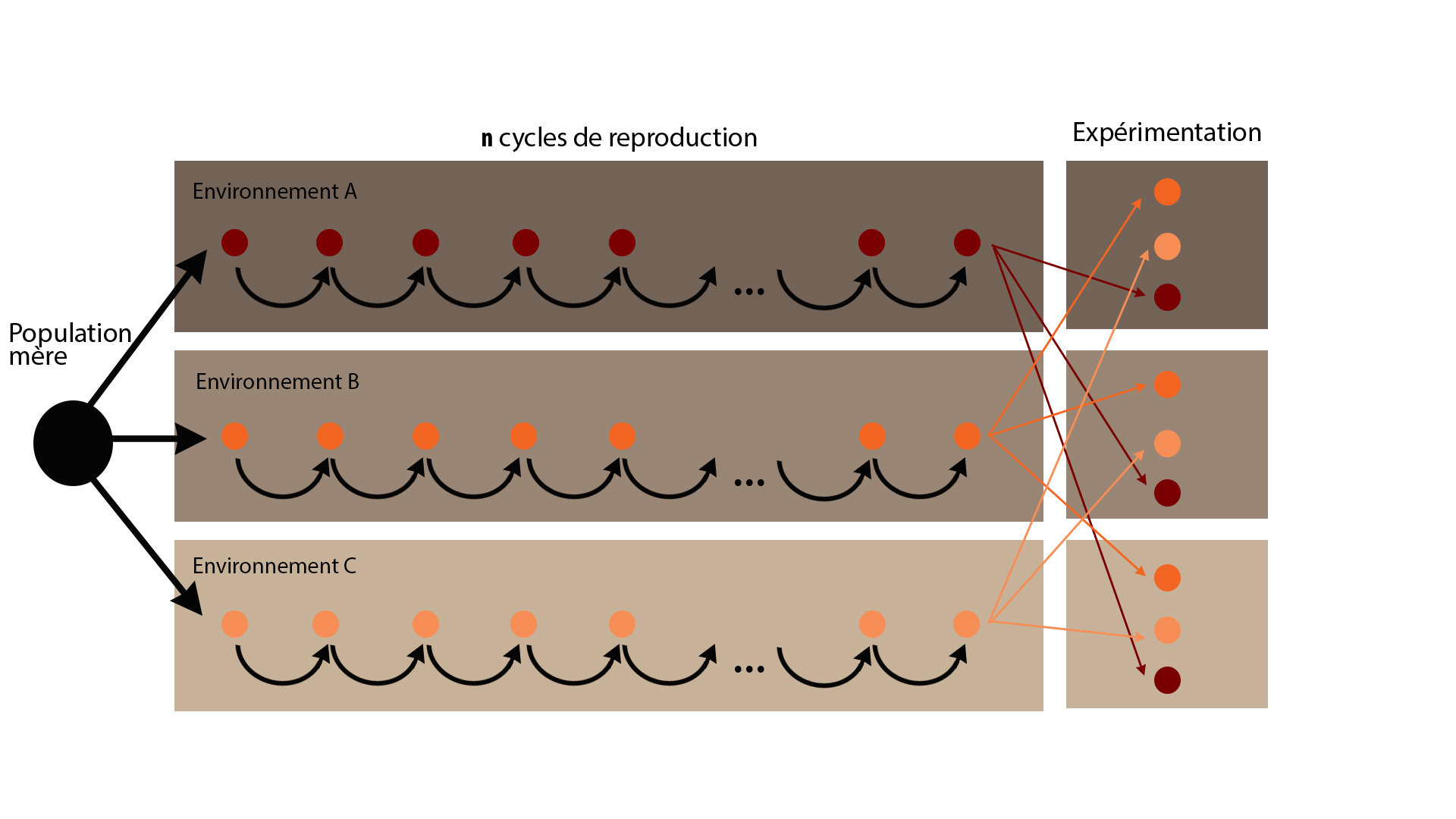
Figure 4: Exemple de protocole expérimental pour évaluer le degré d’adaptation d’une variété à un environnement
Plusieurs études montrent l’absence d’interactions cultivar x système en faisant varier différents éléments de l’agroécosystème. A l’inverse, d’autres études montrent que certains cultivars sont davantage adaptés à un système plus qu’à un autre. Le tableau (table 1) présente un échantillon d’études, et résultats associés, relatifs à l’adaptation de cultivars à des agroécosystèmes spécifiques.
| Publication | Elément.de.l.agroécosystème.étudié | Espèce.étudiée | Effet.d.interaction.cultivar.x.système |
|---|---|---|---|
| (Carr et al., 2003) [8] | Labour/sans labour | Blé | Non |
| (Rao and Dao, 1994) [9] | Labour/sans labour | Blé | Non |
| (Weisz and Bowman, 1999) [10] | Labour/sans labour | Blé | Non |
| (Panter and Allen, 1989) [11] | Association de culture | Soja | Non |
| (Santalla et al., 2001) [12] | Association de culture | Haricot | Non |
| (Hasegawa, 2002) [13] | Fertilisation azoté | Riz | Non |
| (O’Leary and Smith, 2009) [14] | Association de culture | Maïs | Oui |
| (Atlin and Frey, 1990) [15] | Système bas intrants | Orge | Oui |
| (Brancourt-Hulmel et al., 2005) [16] | Système bas intrants | Blé | Oui |
| (Brun and Dudley, 1989) [17] | Système bas intrants | Maïs | Oui |
| (Ceccarelli, 1994) [18] | Système bas intrants | Orge | Oui |
| (Murphy et al., 2005) [7] | Système bas intrants | Blé | Oui |
Dans la plupart de ces études, le processus de sélection naturelle n’est pas directement étudié. Cependant, les interactions complexes cultivar x environnement mises en évidence permettent de justifier en partie l’effet positif d’un environnement spécifique sur la capacité d’une « génétique » donnée à produire une descendance (germination, survie, production de grain). Le degré d’évolution d’une population sous l’unique effet de la pression de l’agroécosystème (absence de sélection consciente) est très peu étudié dans la littérature.

Figure 5: La pratique du désherbage à la herse étrille sur une population constitue une pression de l’environnement (pratique culturale) sur les plantes et favorise les individus avec un fort enracinement, cette action répétée sur plusieurs générations induit une adaptation génétique de la population à la pratique de l’agriculteur
Ce qu’il faut retenir
La sélection naturelle ou l’adaptation sont des processus très complexes car en interaction avec de nombreux éléments de l’agroécosystème
On peut observer une adaptation des variétés sur une dizaine d’années environ
Plus la pression de l’environnement est forte, plus l’adaptation est rapide
Les études scientifiques sur la sélection naturelle chez les plantes cultivées sont peu nombreuses
Il est plus facile d’étudier le caractère « adapté » d’une variété (ex la variété A se porte mieux que la variété B dans l’environnement 1 et inversement dans l’environnement 2) que le processus d’adaptation.
Les idees recues ou les affirmations encore non verifiees scientifiquement
-“Quand on récupère une variété, il faut laisser la population “s’adapter” au territoire avant de faire de la sélection massale” : cette affirmation n’est pas objectivée scientifiquement, cependant, on peut imaginer que dans certaines conditions extrêmes, le changement d’environnement pour la variété serait tellement fort (exemple changement de continent) que la pression environnementale empêcherait la plupart des individus de produire une descendance viable, et que dans ce cas, on peut faire le choix de conserver toute la génétique qui est capable de survivre et qui s’est recombinée (pour les plantes allogames comme le maïs ou le tournesol) avant d’effectuer une sélection consciente. Mais dans une situation où la variété bouge de quelques centaines de km, même si les systèmes agricoles sont différents, nous conseillons de ne pas perdre de temps et de commencer dès la première année à faire de la sélection massale (pour les agriculteurs souhaitant améliorer leur variété).
-“On peut compter uniquement sur la sélection naturelle pour que la variété s’améliore” : cette hypothèse est en partie vraie, surtout si la pression environnementale est très forte (ou conditions culturales inappropriées à l’espèce cultivée) (exemple de la résistance du soja aux nématodes). Cependant, la sélection naturelle est, dans la très grande majorité des cas, un processus très lent. Il faut une dizaine d’années environ pour observer une différence statistiquement significative mais il faut beaucoup plus de temps pour que l’amélioration soit agronomiquement intéressante. Ce n’est pas en cultivant des tomates deux ans sans eau que vous développerez une variété résistante à la sécheresse. Certains traits cependant évoluent naturellement plus vite que d’autres, comme la précocité de la plante [19].
-“La population garde le souvenir d’un épisode de stress et le transmet à la génération suivante”. Ces affirmations viennent de découvertes génétiques récentes et d’un nouveau champ de connaissances que l’on nomme “épigénétique” et qui est souvent résumé (mais un peu abusivement) comme “l’héritabilité des caractères acquis”. Pour le moment, on ne connait que très peu de choses sur ces mécanismes (un article y sera consacré). Les quelques effets de l’épigénétique aujourd’hui identifiés sont que, dans certaines conditions (méconnues), les descendants sur 2 ou 3 générations d’une population qui a subi un stress (famine sur des populations humaines) présentent une espérance de vie plus courte. Du point de vue de la sélection paysanne, il est encore impossible de dire si ces mécanisme sont positifs ou négatifs et s’ils peuvent être prédis, voire pilotés. Pour mieux comprendre l’épigénétique, vous pouvez visionner une très bonne vidéo de vulgarisation de la chaîne “science étonnante” ici.
Sources
[1] S. Jain, “Studies on the Breeding of Self-Pollinated Cereals,” Euphytica, vol. 10, pp. 315–324, 1961.
[2] A. Wezel, S. Bellon, T. Doré, C. Francis, D. Vallod, and C. David, “Agroecology as a Science, a Movement and a Practice. A Review,” Agronomy for sustainable development, vol. 29, pp. 503–515, 2009.
[3] M. Sebillotte, “Agronomie et Agriculture. Essai d’analyse Des Tâches de l’agronome,” Cahier ORSTOM - Série biologie, no. 24, pp. 3–25, 1974.
[4] J. Enjalbert, I. Goldringer, S. Paillard, and P. Brabant, “Molecular markers to study genetic drift and selection in wheat populations,” Journal of Experimental Botany, vol. 50, no. 332, pp. 283–290, 1999.
[5] K. M. Ibrahim, J. Hayter, and J. Barrett, “Frequency changes in storage protein genes in a hybrid bulk population of barley,” Heridity, vol. 77, no. 3, pp. 231–239, 1996.
[6] V. Le Boulc’h, J. David, P. Brabant, and C. Vallavieille-Pope, “Dynamic conservation of variability: Responses of wheat populations to different selective forces including powdery mildew,” Genetics selection evolution, vol. 26, no. 1, pp. 221–240, 1994.
[7] K. Murphy, D. Lammer, S. Lyon, C. Brady, and S. S.Jones, “Breeding for Organic and Low-input Farming Systems: An Evolutionary–participatory Breeding Method for Inbred Cereal Grains,” Renewable Agriculture and Food Systems, vol. 20, no. 1, pp. 48–55, Mar. 2005.
[8] P. M. Carr, R. D. Horsley, and W. Poland, “Tillage and Seeding Rate Effects on Wheat Cultivars,” Crop Science, vol. 43, no. 1, p. 202, 2003.
[9] S. C. Rao and T. H. Dao, “Straw Quality of 10 Wheat Cultivars under Conventional and No-Till Systems,” Agronomy journal, vol. 86, no. 5, p. 833, 1994.
[10] R. Weisz and D. T. Bowman, “Influence of Tillage System on Soft Red Winter Wheat Cultivar Selection,” Journal of Production Agriculture, vol. 12, no. 3, pp. 415–418, 1999.
[11] D. Panter and F. Allen, “Simulated selection for superior yielding soybean lines in conventional vs double-crop nursery environments,” Crop Science, vol. 29, no. 6, 1989.
[12] M. Santalla, P. Rodino, P. Casquero, and A. M. de Ron, “Interactions of Bush Bean Intercropped with Field and Sweet Maize,” European Journal of Agronomy, vol. 15, no. 3, pp. 185–196, 2001.
[13] H. Hasegawa, “High-yielding rice cultivars perform best even at reduced nitrogen fertilzer rate,” Crop Science, vol. 43, no. 3, pp. 921–926, 2002.
[14] N. O’Leary and M. Smith, “Breeding corn for adaptation to two diverse intercropping companions,” American Journal of Alternative Agriculture, vol. 14, no. 4, pp. 158–164, 2009.
[15] G. Atlin and K. Frey, “Selecting Oat Lines for Yield in Low-Productivity Environments,” Crop Science, vol. 30, no. 3, 1990.
[16] M. Brancourt-Hulmel et al., “Indirect versus Direct Selection of Winter Wheat for Low-Input or High-Input Levels,” Crop Science, vol. 45, no. 4, 2005.
[17] E. Brun and J. Dudley, “Breeding Potential in the USA and Argentina of Corn Populations Containing Differnet Proportions of Flint and Denbt Germplasm,” Crop Science, vol. 29, no. 3, 1989.
[18] S. Ceccarelli, “Specific Adaptation and Breeding for Marginal Conditions,” Euphytica, vol. 77, no. 3, pp. 205–219, 1994.
[19] E. Serpolay, S. Giuliano, N. Schermann, and V. Chable, “Evaluation of Evolution and Diversity of Maize Open-Pollinated Varieties Cultivated under Contrasted Environmental and Farmers’ Selection Pressures: A Phenotypical Approach,” Open Journal of Genetics, vol. 4, no. 2, pp. 125–145, Apr. 2014.